A legnépszerűbb, leggyakoribb és tudományosan legalaposabban feldolgozott fosszilis állatcsoport, mégis rengeteg dolgot nem tudunk róluk. Pontosan hogyan néztek ki? Mit ettek? Hogyan mozogtak? Ezek a kérdések a mai napig heves vitákat szülnek a paleobiológia kutatóinak körében. Megpróbáltam utánajárni, hogy áll ma a tudomány a válaszokkal kapcsolatban.
A fejlábúak Ammonoidea alosztálya nagyjából 400 millió évvel ezelőtt tűnt fel az élet színpadán, és bár törzsfejlődésük első szakaszában, az állati óidő végéig alárendelt szerepet játszottak, a perm végi nagy kihalást követően egészen a kréta időszak legvégéig uralták a tengeri életközösségeket.
Az általános szóhasználatban ammonitesz gyűjtőnévvel ellátott állatok aragonitból kiválasztott külső vázuknak köszönhetően az átlagosnál nagyobb eséllyel fosszilizálódtak, így a mezozoós tengeri karbonátos kőzetek leggyakoribb ősmaradványai közé tartoznak. Mindemellett kozmopoliták voltak, tehát az egész Földön elterjedtek, külső vázuk jól elkülöníthető bélyegekkel rendelkezett, és rendkívül gyors volt a törzsfejlődésük, így az olyan külső jegyek, mint pl. bordák, gumók és egyéb vázelemek, rövid időn belül változtak. Ennél nem is kell több egy rétegtannal, a kőzetek korának meghatározásával foglalkozó szakember számára. Az ammoniták fő tudományos értéke abban rejlik, hogy segítségükkel koruk kőzetrétegei szintekre, ezáltal korszakokra tagolhatóak, elképesztő diverzitásuknak és rövid fajöltőjüknek hála nagyon megbízható módon, akár százezer éves pontossággal. Ez geológiai távlatokat tekintve kiemelkedően precíz kormeghatározást tesz lehetővé.
Az Ammonoideák tehát komoly szerepet töltenek be az alkalmazott tudományok terén, feldolgozottságuk kiemelkedő a fosszíliák sorában. Ám mivel a kormeghatározáshoz a legfontosabb információt a külső váz bélyegei nyújtják, és nem mellesleg ezen kívül az esetek döntő részében más nem is marad fenn az állatból, a tudomány szinte kizárólag ezekkel az alaktani kérdésekkel foglalkozik. A cél ilyenkor elsősorban az ammonitesz példány nemzetség és faj szintű pontos meghatározása, esetleg a paleobiogeográfiai következtetések levonása (az egykori életformák födrajzi elterjedésével foglalkozó tudományág).
Pedig evolúcióbiológiai szempontból felbecsületetlen lenne megfejteni egy ennyire sikeres, hosszúéletű, elterjedt és diverz állatcsoport élettani hátterének titkait.
Hogy nézett ki a külső héj által rejtett élőlény? Milyen volt az életmódja? Mit evett? Hogyan mozgott?
 Így nézhettek ki. Vagy nem. A tudomány előrehaladásával talán többet is megtudunk majd ezekről a titokzatos lényekről. Frank Lode ammonitesz ábrázolása a művész DeviantArt oldalán
Így nézhettek ki. Vagy nem. A tudomány előrehaladásával talán többet is megtudunk majd ezekről a titokzatos lényekről. Frank Lode ammonitesz ábrázolása a művész DeviantArt oldalán
Nem egyszerű azon kutatók dolga, akik ezekre a kérdésekre keresik a választ. Annak ellenére, hogy világszerte elterjedt és hosszú időn át fennálló csoport volt, a hiányos leletanyag jelentősen megnehezíti a paleobiológiai vizsgálatokat. Az ammonitákból ugyanis nem sok maradt meg az utókor számára. A puhatestűek szövetei az elhullás után pillanatok alatt az enyészeté lesznek, ráadásul az ammoniteszek teste a szilárd vázban lévő gázok miatt még sokáig lebeghetett a vízben, ami az esetek döntő többségében megakadályozta a lágytest gyors betemetődését. De még maga a szilárd váz is szinte minden esetben feloldódott még a fosszilizáció előtt, a tengervíz által kifejtett kémiai hatások miatt. Az, hogy mégis látunk ma belőlük bármit is – ez rendszerint a héj úgynevezett kőbele – annak köszönhető, hogy ez a kioldódás lassabban ment végbe, így mire teljesen eltűnt, a bezáró kőzet már volt olyan szilárdságú, hogy egy minimális légréteg maradjon az egykori váz helyén. Persze kivételes lelőhelyeken található szép számmal olyan példány is, amelynek a váza nem oldódott ki, hanem annak anyaga kicserélődött különféle kristályokra, általában kalcitra. Ezek a héjas példányok már nyújtanak némi segítséget a kutatóknak ahhoz, hogy következtetéseket vonjanak le a házban élő állat anatómiai jegyeire, de jelentősen bonyolítja az ügyet az a tény, hogy ezeknél az állatoknál nem voltak annyira egyértelmű kapcsolódási, ízületi pontok a szilárd vázhoz, mint ahogy azt tapasztaljuk az olyan bonyolultabb szervezeteknél, mint a gerincesek vagy az ízeltlábúak. (Az viszont például már a külső váz vizsgálatánál látszik, hogy az ammoniteszek puhateste sokban különbözhetett az egyetlen mai összehasonlítási alapot nyújtó távoli rokonukétól, a nautilusokétól: hosszabb és keskenyebb lakókamrájuk arra utal, hogy a benne élő állat – szerveivel együtt – hosszabb, nyúlánkabb lehetett.)
A fennmaradó szilárd váznál is sokkal, de sokkal ritkábban pedig előfordul, hogy az állat egykori lágytestére utaló nyomok maradnak fenn a kőzetekben. Ezen nyomok egy része szó szerint csupán nyom, ahol a kőzetben az állat élete, vagy esetleg a haláltusája során otthagyott lenyomatok hagytak üzenetet az utókor számára. Azonban több olyan tanulmány is napvilágot látott, amelyben bizonyos nagyon szerencsés körülmények között fosszilizálódott lágyszövetekről írnak. Szerencsére a technológia is egyre hatékonyabb segítséget tud nyújtani a kutatásokhoz: korábban röntgen, újabban CT és más, bonyolultabb eljárásokkal, mint például részecskegyorsítóval (szinkrotonnal) végzett eljárásokkal nyernek új információkat a lágyszövetek formájáról és elhelyezkedéséről.
Az őslénytani munka eredményéhez sok esetben járul hozzá egy másik vizsgálati módszer, az aktualizmus elve. Ez a módszer összefüggéseket keres az egykori és a ma is élő életformák között, ilyen módon a jelen ismert tényei alapján következtet a múltra. Mivel az Ammonoideák egy tejesen, és már nagyon régóta kihalt csoport, ez a vizsgálati módszer enyhén szólva korlátozottan áll a kutatók rendelkezésére.
Az Ammonoideák paleobiológiájának kutatása az utóbbi években – főként néhány új lelet előkerülése és vizsgálati módszer bevezetése miatt – új lendületet vett, heves vitákat váltva ki a téma szakértői között.
Lássuk, mi az, amit ma tudunk róluk, és mi az, ami bizonytalan. Elárulom, az utóbbiból van a több.
Az eltűnt lágytest nyomában.
A perdöntő bizonyítékok csaknem teljes hiánya miatt az Ammonoideák lágytestének anatómiája a legvitatottabb terület. Pedig az olyan kérdések, mint hogy hány karjuk vagy kopoltyújuk volt, volt-e tintazacskójuk, mekkora és milyen látószerveik voltak, stb. jelentősen közelebb vinne minket a csoport eredettörténetének megfejtéséhez. A gyér leletanyag mellett tovább nehezíti a tudományos munkát az is, hogy ha találnak is valamilyen átkristályosodott lágyszövetet egy fosszíliában, annak pontos meghatározása igen nehéz. Vannak olyan szervek, mint például az állkapocs vagy a radula (reszelőszerű rágók), amelyek eleve keményebb anyagból, pl. kitinből álltak, így egy szerencsés kövület esetén ezek még könnyebben felismerhetők. De az olyan, igazán puha szöveteket, mint pl. a köpeny vagy a karok, a legkivételesebb esetben is nehéz felismerni. Érdekesség, hogy az ilyen szervek beazonosításához még aktuálpaleontológiai vizsgálatokat is végeztek: megnézték, hogy a mai belsővázas fejlábúak egyes szervei mennyi idő alatt lesznek az enyészeté (rothadnak el), és az ez alapján felállított időbeli sorrend segített kizárni bizonyos testrészeket a kövületek tanulmányozásánál.
Próbáltam áttekinteni, milyen ismeretanyag áll rendelkezésünkre a lágyszöveteket is tartalmazó Ammonoidea fosszíliákkal kapcsolatban, ehhez pedig nagy segítséget nyújtott a téma egyik svájci szakértője, Christian Klug és csapatának munkája. Szerencsére nem is olyan régen, 2015-ben adtak ki egy összegző művet Ammonoid Paleobiology: From Anatomy to Ecology címmel, amelyben egy egész fejezet foglalkozik az ammoniteszek lágytestének anatómiájával, számba véve az eddig előkerült kivételes példányokat.
Klug munkájából kitűnik, hogy a beazonosított szervek a kövületek döntő többségénél a fent is említett rágószervekhez tartoznak, mivel ezek főként kitinből álltak, jobb eséllyel maradnak fenn. Utána következnek az emésztőszervek (gyomor és bélrendszer), kisebb számú példánnyal a nyelőcső, a kopoltyú és a köpeny. Sajnos az olyan fontos szervek, mint a szemek vagy a karok/csápok maradványát, csak feltételezik néhány fosszíliánál.
 Baculitid ammonitesz a cenomán és turon korszakok határáról észak-nyugat Németországból. Az utóbbi idők egyik legizgalmasabb, beazonosítható Ammonoidea lágyszöveteket is tartalmazó fosszíliája (Klug et al, 2012)
Baculitid ammonitesz a cenomán és turon korszakok határáról észak-nyugat Németországból. Az utóbbi idők egyik legizgalmasabb, beazonosítható Ammonoidea lágyszöveteket is tartalmazó fosszíliája (Klug et al, 2012)
Továbbá heves vita zajlott az elmúlt évtizedekben arról, hogy lehetett-e az ammoniteszeknek – vagy legalábbis egy csoportjuknak – tintazacskója. Néhány fosszíliánál bizonyos elszenesedett részek alakja és elhelyezkedése alapján azt feltételezték, hogy az adott szerv tintazacskó lehetett, majd ugyanez a kutatócsoport pár évvel később visszavonta az eredményét. Más kutatók később hasonló maradványokból melanint mutattak ki, ami hasonló következtetéseket szült, de aztán későbbi vizsgálatok bebizonyították, hogy a fejlábúak más szervei is hozhatnak létre melanint, így a tintazacskó jelenlétét ismét nem sikerült hitelt érdemlően bizonyítani.
Az alábbi táblázat a téma ismert kutatói, Christian Klug és Jens Lehmann fent is említett 2015-ös összefoglaló munkájából való, és az addig valaha nyílvánosságra hozott Ammonoidea lágyszövet maradványokat rendszerezi.
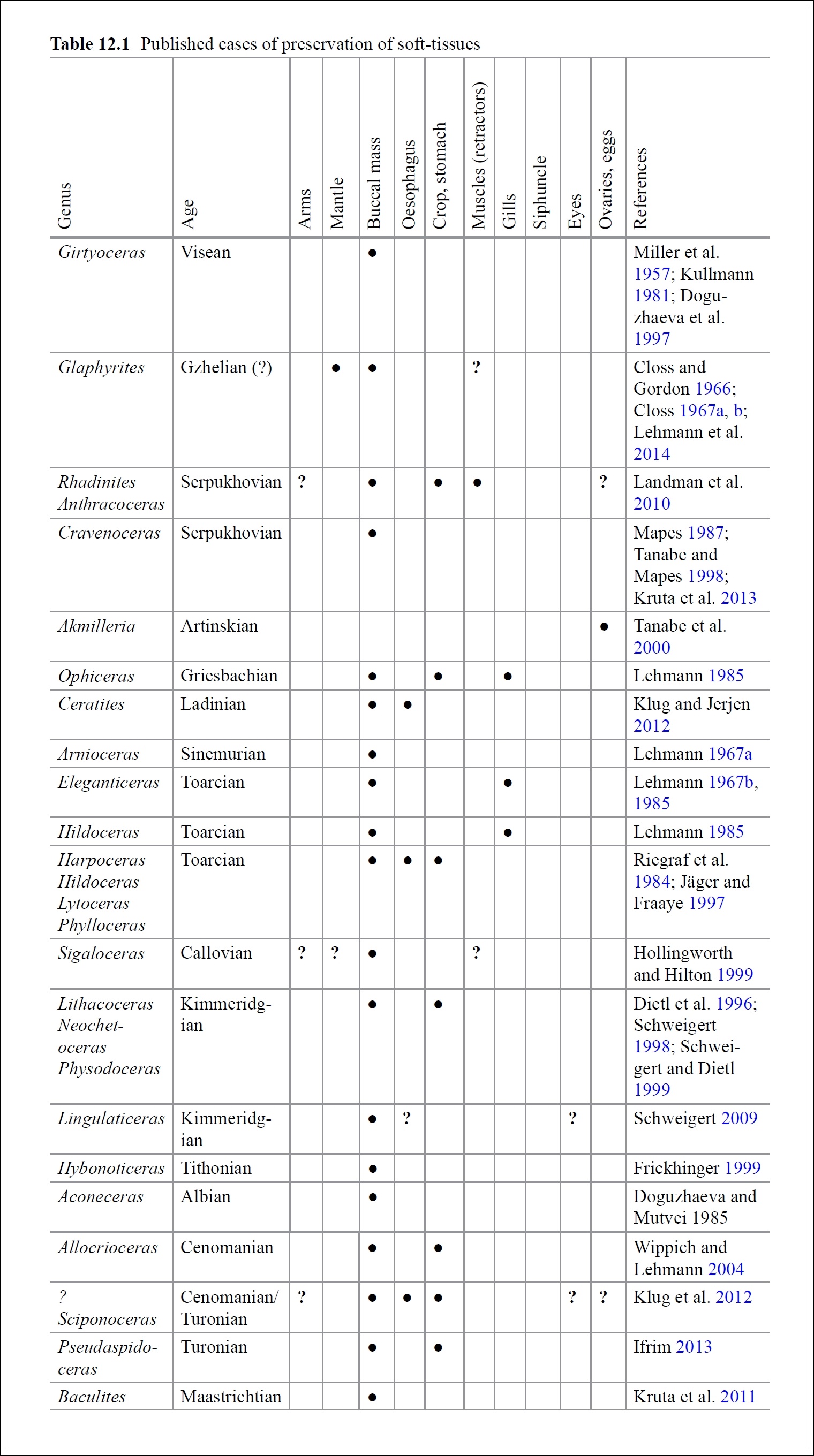
KARdinális kérdés
Biztos vagyok benne, hogy lágyszövet-ügyben a legizgalmasabb kérdés, hogy hány karja lehetett az ammoniteszeknek? Mivel minden ma élő külső és belsővázas fejlábúnak vannak karjai, vagy ha úgy tetszik csápjai, amelyek a szájszerv körül helyezkednek el, nagyon valószínű, hogy az ammoniteszeknek is voltak ilyen szervei. A mai tintahalaknak és polipoknak 8-10 tapogatója van, a nautiluszoknak akár 90 is lehet. Sajnos az eddigi fosszíliarekord azon néhány darabja, ahol feltételezik, hogy a kőzetlapon látható feketés maszat egykor az állat csápja lehetett, nem tekinthető tárgyi bizonyítéknak. Így marad a spekuláció. A korábbi gondolkodásmód az volt, hogy mivel a nautiluszok külső vázukkal valószínűleg a korai fejlábúak jegyeit viselik magukon, hasonló lehet a helyzet a karok számát tekintve is, és az ammoniteszek is több csáppal rendelkeztek, ez a tulajdonság egyfajta archaikus bélyege a Cephalopodáknak. Időközben azonban több olyan tanulmány is napvilágot látott, ahol az Ammonoideák raduláit (rágó szerveit) vizsgálva egyre inkább arra a következtetésre jutottak, hogy rendszertanilag közelebb állhattak a mai belső vázas fejlábúakhoz. Klug és Lehmann pedig, a már említett 2015-ös tanulmányukban aktuálpaleontológiai vizsgálatokat is közzétettek. Ebben arra jutottak, hogy a mai fejlábúak embrionális fejlődése során az embró korai szakaszaiban a karok száma minden esetben mindössze 10. A nautilus magzata is így kezdi, majd a későbbiekben ez a kevés kar válik szét több kisebb csápra az egyedfejlődés során. Ez arra utal, hogy a kisebb számú kar a primitívebb jegy, így az egyéb, Coleoideákkal való rokonságra utaló jelek mellett a ma kijelenthető legvalószínűbb feltételezés az, hogy az ammoniteszeknek kisebb számú karja lehetett, a legvalószínűbb, hogy 8 vagy 10. Ezt a felvetést egyébként tovább erősíti néhány paleozoós nyomfosszília, amelyek szintén kevesebb végtagra utalnak.
Hozzáteszem, az ammoniteszek a földtörténet egyik leghosszabb ideig fennálló csoportja, közel 350 millió éves „pályafutásuk” alatt bőven lehetett idejük száz és százféle alakká fejlődni.
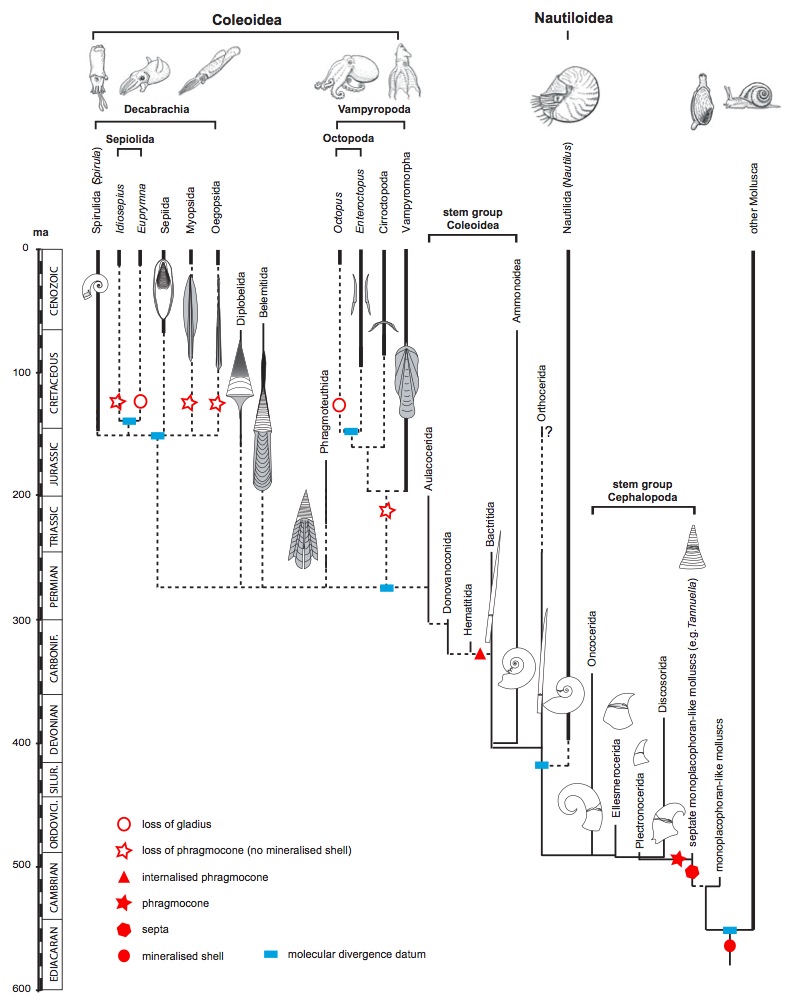
Annak ellenére, hogy a lágyszövetek maradványai ilyen korlátozottan állnak rendelkezésünkre, a tudomány korábbi következtetéseit megerősitik: az Ammonoideákat rendszertanilag továbbra is a puhatestűek törzsén belül a Cephalopoda, azaz a fejlábúak osztályába soroljuk, és több eredmény sugallja azt, hogy anatómiailag, így valószínűleg filogenetikailag is közelebb álltak a mai belső vázas fejlábúakhoz (Coleoidea), mint a Nautiloideákhoz. Azonban hogy pontosan milyen volt a testfelépítésük, és ebből a szempontból melyik ma is élő csoporthoz hasonlítottak leginkább, erősen kétséges.
Egy német-amerikai kutatócsoport 2011-ben tanulmányt adott ki a cephalopodák feltételezett eredettörténetéről és evolúciójáról. Ebben a munkában segítségül hívták a molekuláris evolúcióbiológia technológiai vívmányait is, és a molekuláris óra eljárásával térképezték fel a jelenleg élő fejlábúak filogenetikáját. Nagyon érdekes, hogy ez a vizsgálat minden eddigi, fosszíliák alapján kialakított következtetést megerősített, és az Ammonoideák csoportjának megjelenését is az eddig feltételezett időszakra, a kora devonra teszi. Ami a családfát illeti, a molekuláris vizsgálatok azt is megerősítették, hogy az Ammonoideák a mai belső vázas fejlábúak vonaláról, egy a nautiluszoktól már korábban elkülönült ágról, a Bactritida rendről váltak le. Ez további hitelt ad azoknak a feltételezéseknek, amelyek az Ammoniteszek anatómiai jegyeit a mai belsővázas fejlábúakéval analogizálják.
Napi menü
Jóval egzaktabb a helyzet az Ammonoideák táplálkozásával kapcsolatban, egész tisztán kivehető fosszíliák állnak a rendelkezésünkre annak kapcsán, hogy miből állt a néhai csigaházas polipok étrendje. Ezek a leletek fontos információkkal szolgálnak a paleoökológia számára, nem csak arról, hogy mi volt az ammoniteszek helye a táplálékláncban, de arról is, hogy melyik tengeri élettér elemei voltak. Szintén Klug és Lehmann műve az, amiben részletes elemzést olvashatunk az eddig leírt ammonitesz táplálékot is tartalmazó kövületekről. Az itt bemutatott eredmények alapján az ammonoideák táplálékai foraminiferák (szilárd vázú egysejtűek), planktonikus tengeri liliomok, apró rákok, csigák, kagylók és szivacsok lehettek. Ezek a következtetések mind olyan fosszíliák alapján történtek, amelyeknél az ammonitesz héj lakókamrájában egyéb maradványokat is kimutattak. Ezen példányok néhányára vannak alternatív magyarázatok is: ahol kisebb méretű ammonoidea héjak vannak, ott meg kell vizsgálni, hogy nem embrionális maradványokról van-e szó, esetleg elképzelhető, hogy az állat saját testében gondozta az ivadékát. Más töredékes vázmaradványoknál pedig az is előfordulhat, hogy azok utólag, a betemetődés során sodródhattak a megüresedett lakókamrába. Ettől függetlenül vannak olyan leletek, ahol egyértelmű, hogy az állat elfogyasztott táplálékáról van szó, ezek a maradványok pedig egytől egyig arra utalnak, hogy az Ammonoideák mikropredátorok, vagy más szóval mikrofág életmódot folytató állatok voltak, azaz kisméretű élőlényekkel táplálkoztak. Ezen életmód feltételezése egyébként összecseng azokkal a vizsgálatokkal, amelyek az ammoniteszek rágószerveit vizsgálta. Ez arra a következtetésre jutott, hogy a mezozoós formák többsége képtelen lehetett nagyobb méretű táplálék megragadására és elfogyasztására. Maradtak tehát a mikroszkopikus méretű planktonikus lények, esetleg kisebb úszó vagy aljzathoz rögzített puhatestűek, tüskésbőrűek. Arra azonban jelenleg nincs egyértelmű adat, hogy mekkora részük szerezte be ezt a táplálékot úszva, vagy a vízben lebegve, és mekkora részük az aljzatról felcsipegetve az ott élő, vagy elhullott állatokat.
A planktonikus tengeri liliomok talán legismertebb példája a Saccocoma, amely a híres solnhofeni litográf pala egyik leggyakoribb ősmaradványa, és több alkalommal találták meg vázelemeiket ammonitesz fosszíliák lakókamrájában.
 Saccocoma - sokuk végezte az ammoniteszek gyomrában.
Saccocoma - sokuk végezte az ammoniteszek gyomrában.
További érdekes, de bizonyításra eddig nem került felvetés a Lumbricaria esete, amely a solnhofeni felső-jura litográf pala egy nyomfosszíliája, egészen pontosan koprolitként, azaz fosszilis ürülékként azonosították. Mivel ez a leggyakoribb koprolit típus ebben a formációban, és az ammoniteszek a leggyakoribb testfosszíliák (továbbá a méret is stimmel), elképzelhető, de szigorúan csak elképzelés, hogy a Lumbricaria az Ammonoideák megkövült ürüléke lehet.

Egy figyelemre méltó ammonitesz ábrázolás egy DeviantArt művésztől. Ha elfogadjuk azt az eddig talán legszilárdabb talajon álló feltételezést, hogy az amoniteszek mikropredátorok voltak (ld. fent), valószínűbb, hogy karjaik inkább ilyenek lehettek. A művész a robusztus, polipszerű karok helyett egy a táplálék elragadását segítő, serteszerű képlet vizionált.
Szájfedő vagy nem?
Van azonban olyan kétely is, amely nem is a lágyszövetek erősen hiányos leletanyagával kapcsolatos, hanem meglepő módon a szilárd váz egyik eleméhez köthető. Az ammoniták egyik vázeleme a kagylóhéjra emlékeztető aptychus, amely egy még az aragonitnál is ellenállóbb anyagból, kitines kalcitból épült fel, így még akkor is megmaradt, amikor az állat háza már rég feloldódott. Ezért fordulhat elő, hogy bizonyos kőzetek kizárólag aptychusokat tartalmaznak. Mivel a végleges betemetődés előtt a vázelemek szinte mindig szétestek, az aptychusokat rendszerint az ammoniteszek vázától teljesen elkülönülten találjuk meg. Emiatt sokszor azt sem tudjuk, melyik aptychus melyik ammonitesz nemhez tartozik. És azt sem, hogy pontosan mire használták ezt a primitívebb formáknál egyetlen, fejlettebbeknél két egyforma, tükörszimmetrikus elemből álló szervet. A nautilusoknál van egy ehhez hasonló – bár csupán szaruból álló – vázelem, amelyet egyfajta lakókamrát védő elemként használnak, és néhány kutatás arra az eredményre jutott, hogy ahol be tudták azonosítani „tulajdonosát”, ott az aptychus mérete megegyezett a gazda szájadékának méretével. Ebből arra következtettek, hogy ezek egyfajta szájfedők lehettek, amelyeket a lakókamra nyílására tudtak húzni az állatok, ha védelemre volt szükségük. Azonban ez a magyarázat a beazonosítható leletek többségénél nem állta meg a helyét, a legtöbb esetben ez a méretazonosság nem jelentkezett.
 Neochetoceras példánya Solnhofenből, eredeti helyzetében fosszilizálódott aptychussal.
Neochetoceras példánya Solnhofenből, eredeti helyzetében fosszilizálódott aptychussal.
Ulrich Lehmann és Nicol Morton voltak az elsők, akik néhány élethelyzetben megőrződött fosszília alapján egy alternatív magyarázattal álltak elő: szerintük az aptychus az ammoniteszek állkapcsának része volt. Egy olyan csőrszerű rágószervet feltételeztek, mint amilyen a mai polipoknak is van, és az aptychus ebben a képletben az állkapocs alsó felét képezi. Hipotézisük szerint az ammoniteszek az aptychust egyfajta kanálként használva, azt az aljzat üledékébe mélyesztve szedték össze táplálékukat, amelyek főként foraminiferák és rákfélék voltak – ahogyan azt az olyan fosszíliák is igazolják, amelyeknél egyértelműen be tudták azonosítani néhány Ammonoidea béltartalmát.
A szakma kétkedve fogadta ezt az elméletet, mert mindamellett, hogy „túlgondolt” ötletnek tartották, az aptychusok a gazdaállathoz mérten gyanúsan nagyméretű állkapcsot jelentenének, egy olyan méretű szájszervet, amely indokolatlan lett volna a magyarázatban szereplő művelethez. És mint tudjuk, a természet nem pazarol az erőforrásokkal. Idegenül hangzott továbbá az a feltételezés is, hogy ez az ammoniteszek többségénél aljzathoz kötött, azaz bentikus életmódot jelentett volna. Pedig más tényezők is alátámasztani látszanak ezt a feltételezést. Lássuk, melyek ezek!
Tudtak egyáltalán lebegni?
Hogyne tudtak volna? – végjuk rá első szuszra. Hiszen ez az egyik legegyértelműbben kikövetkeztethető tulajdonságuk: a szilárd vázuk fennmaradt, így tudjuk, hogy hasonló felépítéssel rendelkeztek, mint a nautilusok. Házuk kamrákra volt osztva, amelyet egy vékony csőszerú képződmény, a szifó kötött össze, ennek segítségével szabályozta az állat a kamrákban lévő anyagok, a tengervíz és a gáz arányát, mint ahogy a mai nautilusok is teszik. Ezzel a mechanikai eszköztárral volt képes az állat a fajsúlyát, és ezáltal a vízoszlopban elfoglalt helyét változtatni. Ahogy nőtt az állat, úgy épített hozzá a vázához mindig új meg új, nagyobb méretű kamrákat, ezáltal fenntartva a lebegés lehetőségét, elkerülve, hogy a teste túlnőjön a gáz térfogatából adódó felhajtóerő nagyságán.
Csakhogy van egy kis bökkenő. Egy német mérnök, Klaus Ebel ugyanis kicsit alaposabban körüljárta a dolgot, és kísérleti módszerekkel arra a következtetésre jutott, hogy az ammoniteszek nem voltak alkalmasak a lebegésre. Ahhoz ugyanis, hogy egy ilyen állat váza és puhateste megfelelő felhajtóerőt generáljon, a saját testénél nagyobb tömegű vízet kell, hogy képes legyen kiszorítani a házából. A nautilusok ezt a feladatot csont nélkül elvégzik, hiszen relatíve kisméretű testükhöz megfelelően nagy üreges váz tartozik. Az ammoniteszek azonban jóval hátrányosabb testtömeg/váztérfogat aránnyal rendelkeztek, ami fizikailag nem tehette lehetővé ezt az életformát. Magyarul nem tudtak annyi gázt tölteni a vázukba, ami megemelte volna a nagy popójukat.
Ebel kísérleteit jura időszaki ammoniteszeken végezte, és ezeket aljzatlakó élőlényekként írja le. Feltételezései szerint vázuk arra ugyan alkalmas volt, hogy a tengerfenékről elrugaszkodva kisebb ugrásokkal manőverezzen, de nektonikus, azaz nyíltvízi, lebegésre képes életmódra aligha. További megerősítést lát abban a tényben, hogy az ammonitesz vázak nagy részének alakja sem volt megfelelő a nektonikus életmódra.
 A többségtől, azaz a spirál alakban feltekeredett vázú ammoniteszektől eltérően a csoport heteromorf tagjai a legelképesztőbb formákkal hozzák zavarba a paleobiológusokat. Milyen környezeti tényező késztethette az ilyen formák létrejöttét? Nipponites (balra) és Diplomoceras (jobbra), Franz Anthony illusztrációja.
A többségtől, azaz a spirál alakban feltekeredett vázú ammoniteszektől eltérően a csoport heteromorf tagjai a legelképesztőbb formákkal hozzák zavarba a paleobiológusokat. Milyen környezeti tényező késztethette az ilyen formák létrejöttét? Nipponites (balra) és Diplomoceras (jobbra), Franz Anthony illusztrációja.
Ez utóbbi felvetéssel más tanulmányok is foglalkoztak, hasonló következtetésre jutva, de vannak érvek, amelyek Ebel felfogása ellen szólnak. Jó példa erre az, hogy ammonitesz vázakat gyakran találnak olyan formációkban, amelyek anoxikus környezetben ülepedtek le. Ilyen körülmények között pedig nem valószínű, hogy aljzathoz kötött életmódot folytatott volna bármilyen élőlény is. Ennek a cáfolatnak a sikerességéhez persze az kell, hogy az ilyen lelőhelyekről előkerült ammonitesz vázak valamelyikéről egyértelműen be tudják bizonyítani, hogy autochton módon ágyazódott be, azaz az élőlény halálát követően a ház szállítódás nélkül temetődött be. A mai nautilusoknál előfordul, hogy az üres házakat a tengeráramlatok többezer kilométerre sodorják, mielőtt a tenger üledéke betemetné azokat!
Annak ellenére, hogy Ebel tanulmányai szilárd matematikai és fizikai alapon nyugszanak, a kutatók többsége ma azt a feltevést fogadja el valószínűbbnek, hogy az ammoniteszek nyíltvízi, mobilis életmódot folytattak, azaz lebegtek.
És mi a helyzet az úszással?
A fentiekben a kamrák gáz-víz arányának változtatásával történő mozgás lehetőségét vettük számba. A mai nautilusok azonban képesek arra is, hogy egy belső szervükbe, az úgynevezett köpenybe felszívják a vízet, és annak kilövellésével, a lökhajtásos elv alkalmazásával gyorsan odébbálljanak, ha valahol kínossá válik a helyzet. A kilövellést magát úgy érik el, hogy köpenyüregük falát összehúzzák, ehhez pedig saját testüket hirtelenjében vissza kell hogy húzzák a lakókamrába. Ezzel tulajdonképpen képesek arra, hogy a nyílt tengeren ide-oda ússzanak, ha kedvük tartja. Vajon az Ammonoideák is képesek voltak ilyesmire?
Nos, mily meglepő, itt is nagy kérdőjelek vannak.
A mai nautilusok lakókamrája ugyanis rövid és széles. Ez épp ideális az olyan hirtelen mozdulatokhoz, mint amilyet a lökhajtásos mozgás érdekében végeznek: egy ilyen lakókamrába a megfelelő köpenyizomzattal könnyedén vissza lehet húzni a testet, hogy kipréselődjön a víz. Az ammoniteszek lakókamrái azonban legtöbb esetben hosszúak és keskenyek voltak – volt olyan nemzetség is, amelynek másfél méter hosszú lakókamrája mindösszesen 3 cm átmérőjű volt! De kevésbé extrém esetben is túl hosszú és keskeny lakókamrával bírtak ahhoz, hogy ezt a mozdulatot ésszerű energia felhasználásával el tudják végezni. Az ezzel kapcsolatos kutatások arra az eredményre vezettek, hogy egy ilyen típusú mozgás sok ammonitesznél egyszerűen azt eredményezte volna, hogy a ház a hirtelen mozdulat következtében minden alkalommal átbukott volna a saját tengelyén. Vicces látvány lenne egy ilyen módon közlekedő tengeri állat, de kétségkívül nehéz elképzelni, hogy ez evolúciós szempontból életképes megoldás lett volna.
Egy másik felvetés egy kicsit az aktualizmus elvéhez nyúl ebben a témakörben. Ha megnézzük a teljes Ammonoidea leletanyagot, ami rendelkezésünkre áll, azt látjuk, hogy a vázak nagy része erősen díszített, amely javarészt bordákat és gumókat jelent. Mindemellett az Ammonoideák kamraválaszfalai a csoport törzsfejlődése során egyre inkább fodrosabbak lettek, ezt egyébként a válaszfalaknak a váz külső részén tanulmányozható lenyomata, az ún. lobavonal mutatja. A külső váz bordái a mai tengeri életközösségekben egyértelműen a bentikus, azaz aljzathoz kötött élőlények bélyegei, de logikus is: ezek a bordák ellenállóbbá teszik a vázat. Ugyanez a helyzet a válaszfalakkal is: minél gyűröttebb, annál nagyobb ellenállást ad az állat házának. Ez duplán megéri annak, aki a tenger mélyén él, nagyobb víznyomás alatt, és ráadásul képtelen gyorsan elinalni egy esetleges támadás elől! A kiváló úszó nautilusok nem rendelkeznek díszítéssel, mert nincs is nagy szükségük rá.
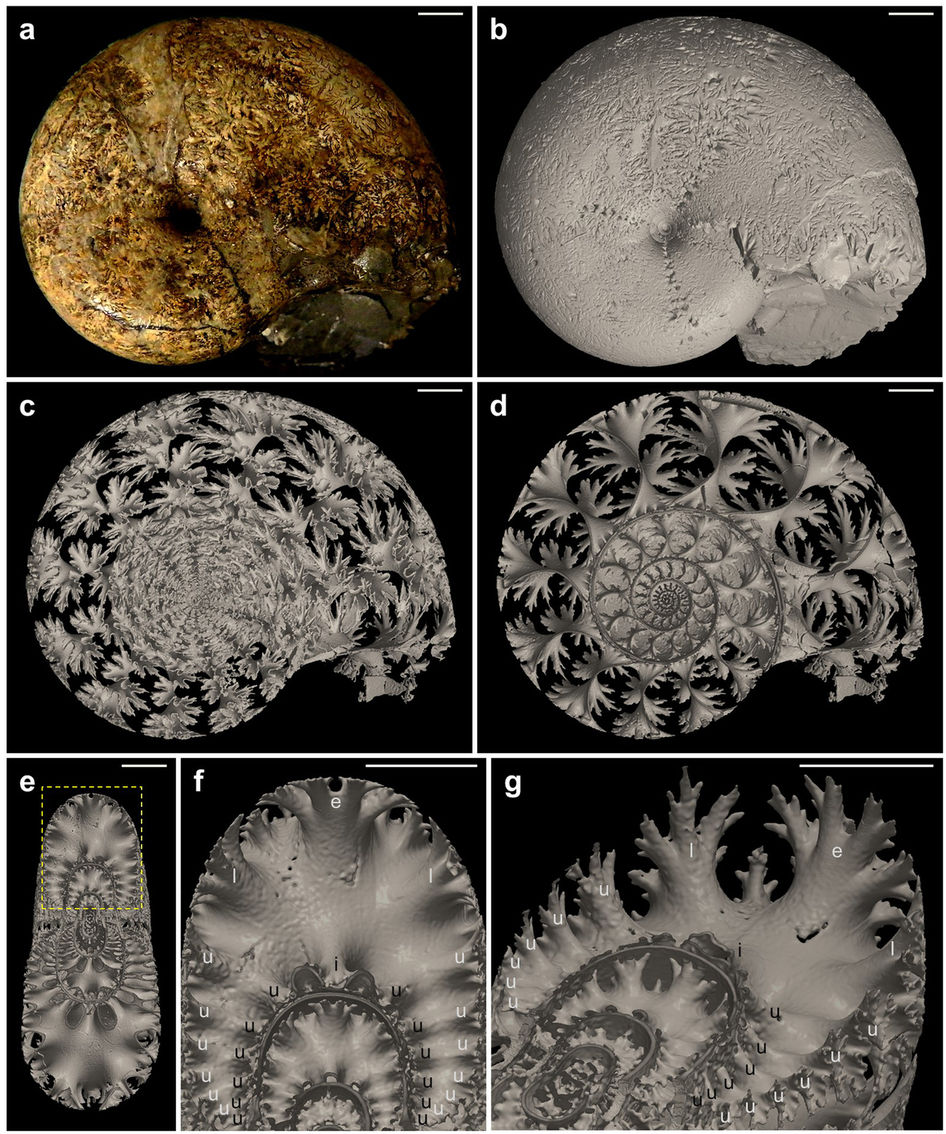 Damesites Damesi kamraválaszfalainak mikro-computer-tomográfiás képe. Az új tecnológiákkal minden eddiginél részletesebben elemezhetők ezek a kulcsfontosságú morfológiai jegyek, amelyek aztán fontos evolúcióbiológiai kérdésekre is választ adhatnak.
Damesites Damesi kamraválaszfalainak mikro-computer-tomográfiás képe. Az új tecnológiákkal minden eddiginél részletesebben elemezhetők ezek a kulcsfontosságú morfológiai jegyek, amelyek aztán fontos evolúcióbiológiai kérdésekre is választ adhatnak.
Elképzelhetőnek tartják, hogy az áramvonalasabb, kevésbé díszített vázú ammoniteszek jobb úszók voltak, míg bordázott társaik nem. Bele sem merek gondolni, mi a helyzet a szerényen díszített, de bonyolult lobavonalakkal megáldott jószágokkal... talán nekik vékonyabb volt a külső vázuk anyaga, ezért volt szükség komolyabb segítségre annak merevítésénél a kamraválaszfalak részéről.
Összegzésként elmondható, hogy a fent említett néhány zavaró tényező ellenére az általános vélemény a szakemberek körében még mindig az, hogy ha nem is mindegyik típus, de az ammoniteszek egy része többé-kevésbé képes volt úszni. Voltak kutatások, ahol az ammonitesz vázak akvadinamikáját tanulmányozták, néhol valós vízárammal egy tartályban, más esetben számítógépes modellekkel (Peter Allison, Imperial College). Ezen vizsgálatok során voltak olyan Ammonoidea csoportok, amelyek váza kifejezetten áramvonalasnak bizonyult, így akár jó úszók is lehettek, mint pl. az Oxynoticeras és a Placenticeras (mellesleg előbbinél minimális, utóbbinál teljesen hiányzik a ház díszítettsége).
A számítógépes kísérletek még érdekesebb eredményt hoztak: volt olyan háztípus, amelynél a házat díszítő bordák nemhogy rontottak volna az áramvonalasságon, hanem még elő is segítették azt.
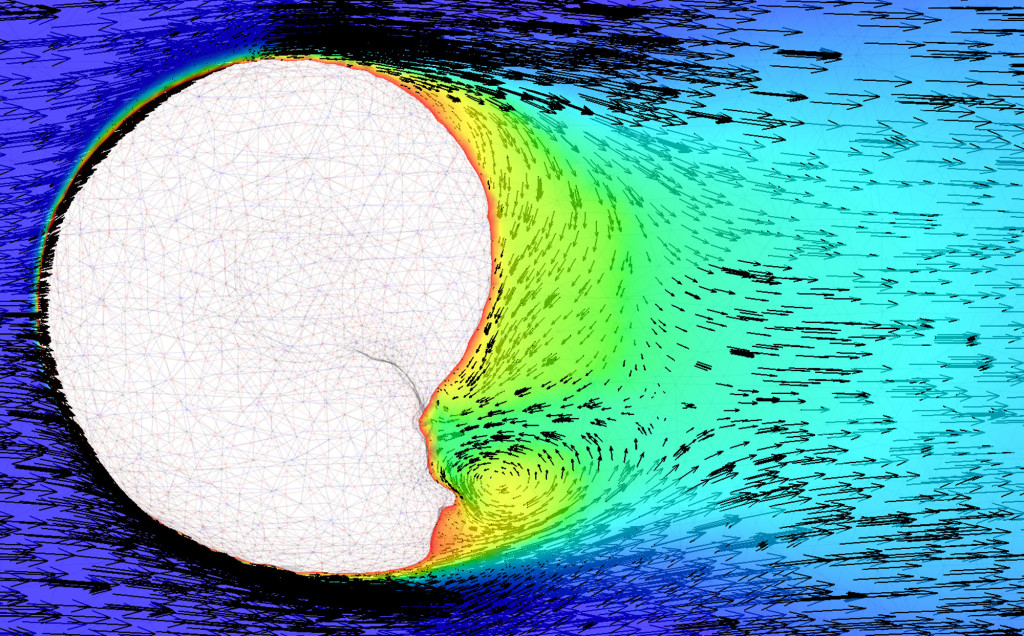 A londoni Imperial College egy kutatócsoportja Dr. Peter Allison vezetésével számítógépes áramlási modellek segítségével tanulmányozta az ammonitesz vázak esetleges úszóképességét.
A londoni Imperial College egy kutatócsoportja Dr. Peter Allison vezetésével számítógépes áramlási modellek segítségével tanulmányozta az ammonitesz vázak esetleges úszóképességét.
Sok nyitott kérdés van tehát az ammonoideák paleobiológiáját illetően, mondhatjuk úgy is, hogy ők a paleontológiai leletanyag ismeretlen ismerősei. A tudomány azonban fejlődik, minden új nap hozhat egy új, ismereteinket bővítő, vagy akár paradigmaváltó fosszíliát. Ugyanígy, a technológia fejlődésével is remélhetőleg újabb módszerek nyitnak majd új távlatokat ebben a kérdéskörben is, és egyre többet tudunk majd ezekről a fontos lényekről.
***
Források:
- Christopher Taylor: The Arms of an Ammonite (Catalogue of Organisms, 2017.02.20.)
- De Baets, K., Hoffmann, R., Sessa, J. A. and Klug, C. 2016. Fossil Focus: Ammonoids. Palaeontology Online,Volume 6, Article 2. 1‐15.
- Deposits Magazine: Ammonite Wars 2016/02/25
- Ebel, K. 1992 04 15: Mode of life and soft body shape of heteromorph ammonites. Lethaia, Vol. 25, pp. 179-193. Oslo. ISSN 0024-1 164.
- Ebel, Klaus. (1990). Swimming abilities of ammonites and limitations. Palsentologische Zeitschrift. 64. 25-37. 10.1007/BF02985919.
- Franz Anthony: 500 million years of cephalopod fossils (Earth Archives 2018 February)
- Géczy Barnabás: Ősállattan - Invertebrata Paleontologia (Nemzeti Tankönyvkiadó, 1993)
- Géczy Barnabás: Őslénytan (Nemzeti Tankönyvkiadó, 1984)
- Inoue S., Kondo S. (2016) Suture pattern formation in ammonites and the unknown rear mantle structure, Scientific Reports, 6 , art. no. 33689
- Klug, Christian & Korn, Dieter & De Baets, Kenneth & Kruta, Isabelle & Mapes, Royal. (2015). Ammonoid Paleobiology: From Anatomy to Ecology. 1007/978-94-017-9630-9.
- KLUG, C., RIEGRAF, W., LEHMANN, J. 2012. Soft–part preservation in heteromorph ammonites from the Cenomanian–Turonian Boundary Event (OAE 2) in north–west Germany. Palaeontology, 55, 6, 1307–1331.
- Kröger, B. , Vinther, J. and Fuchs, D. (2011), Cephalopod origin and evolution: A congruent picture emerging from fossils, development and molecules. Bioessays, 33: 602-613. doi:10.1002/bies.201100001
- Ritterbush, K. A., Hoffmann, R. , Lukeneder, A. and De Baets, K. (2014), Pelagic palaeoecology of ammonoids. J Zool, 292: 229-241. doi:10.1111/jzo.12118
- Robert Lemanis, Stefan Zachow, Florian Fusseis and René Hoffmann (2015). A new approach using high-resolution computed tomography to test the buoyant properties of chambered cephalopod shells. Paleobiology, 41, pp 313-329 doi:10.1017/pab.2014.17



























 A Kínában talált Alalcomenaeus ősmaradvány, a skorpiók és a pókok távoli rokona
A Kínában talált Alalcomenaeus ősmaradvány, a skorpiók és a pókok távoli rokona