Az élővilág történetének egyik sorsdöntő fordulata volt a növények szárazföldi megjelenése és térhódítása, amely elképesztő lendületet adott mind a modern ökoszisztémák kifejlődésének, mind a szárazföldi élővilág kibontakozásához szükséges légköri-éghajlati feltételek kialakulásának. A szárazföldi növénytakaró környezet- és éghajlatformáló tulajdonsága pedig a mai napig hatással van a bolygó minden egyes élőlényére.
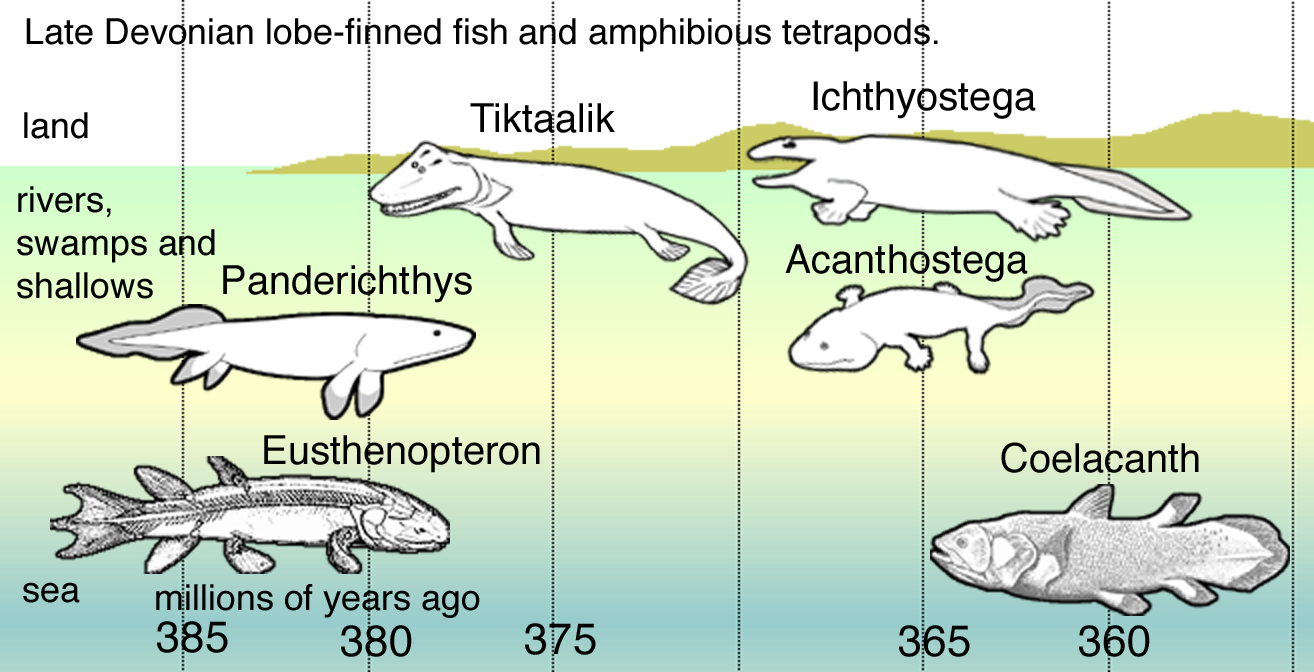
A rendelkezésre álló fosszilis bizonyítékok alapján ma úgy gondoljuk, hogy valamikor az állati óidő szilur időszakának vége felé, nagyjából 430 millió éve kezdték elfoglalni a szövetes, vagy edényes növények - a mai legtöbb növény őseinek első képviselői a tengerekhez közeli, állandó vízborítású területeket, a tavak és folyók környezetét. Makrofosszíliák, azaz szemmel is látható növényi maradványok kb. 430 millió évvel ezelőttig engedik számunkra feltárni a szárazföld növényvilágának családfáját – ma egy Prágában őrzött Cooksonia lelet számít a legősibb növényi makrofosszíliának. Különböző biomarkerek segítségével azonban ennél korábbi kőzetekből is mutattak már ki növényi spórákra utaló jeleket, amely alapján úgy tűnik, hogy már a középső-ordovíciumban – azaz 470 millió éve – megjelenhettek a szárazföldön az első, májmoha-szerű növények.
Meglehetősen idegen látványt nyújthatott ekkoriban a szárazföld, de a késő szilurtól kezdve aztán a fejlettebb harasztok és korpafüvek is egyre nagyobb mennyiségben és változatosságban népesítették be a szárazföldet. Olyannyira, hogy a szilur legvégének kiterjedt növényvilágáról már kőszéntelepek is tanúskodnak. Végül a devon közepére-végére eljutott oda a növényvilág, hogy több fejlődési vonalon is megjelenhettek az nagyobb méretű növények, és kialakulhattak az első, hatalmas fáknak otthont adó erdőségek.
A paleobotanika történetének elmúlt két évszázada során komoly leletanyag halmozódott fel, amely alapján összeállt egy kép a szárazföldi növények korai evolúciójáról és életközösségéről. Van azonban egy fosszilis csoport, ami immár több, mint másfél évszázada borzolja a kedélyeket az ősnövénytan területén.
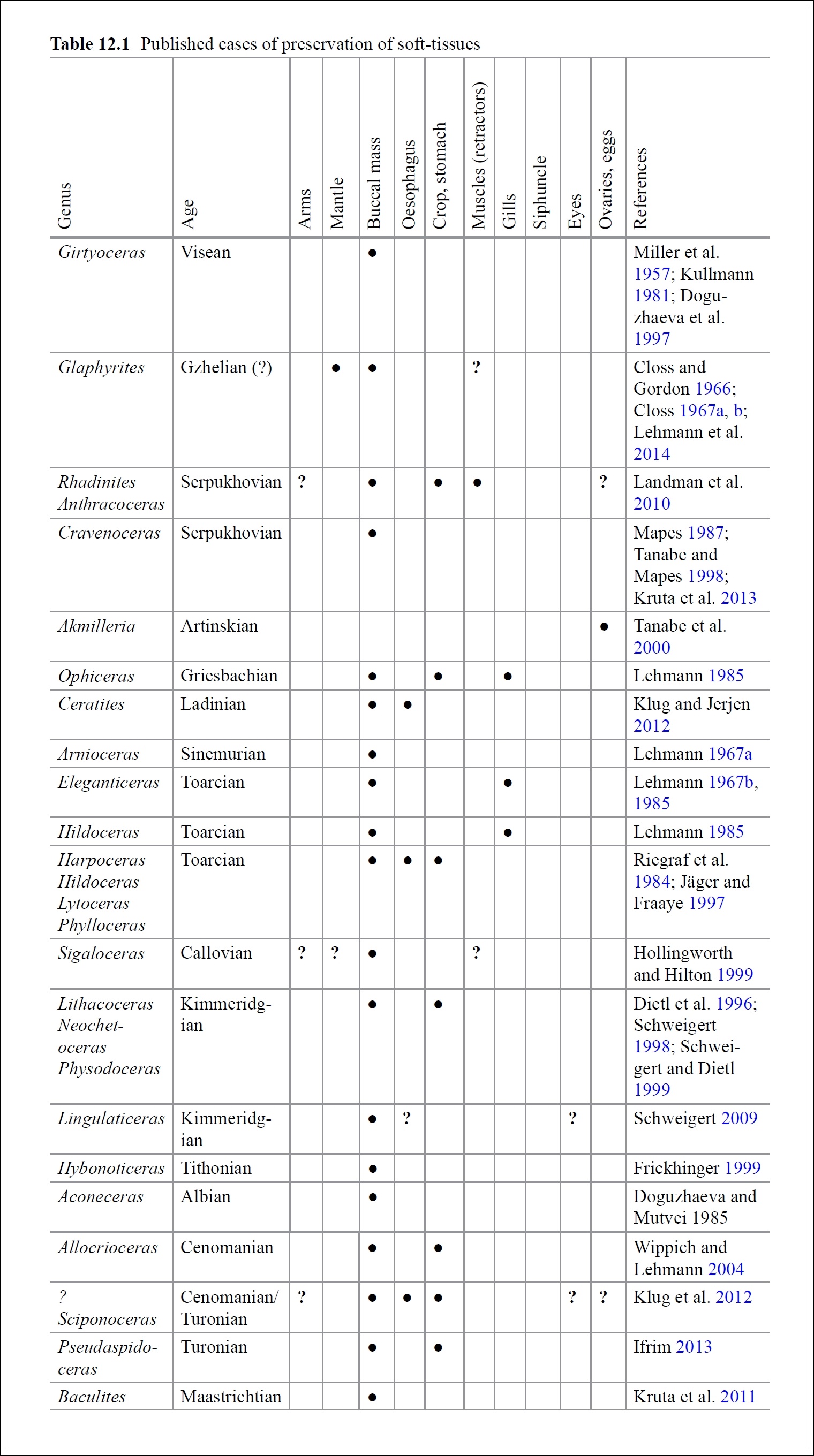
 A szárazföld úttörői - kora-devon látkép, 408 millió éve (illusztráció: Jurij Priymak)
A szárazföld úttörői - kora-devon látkép, 408 millió éve (illusztráció: Jurij Priymak)
Egy igazi óriásról van szó, amely már akkor nyolc méter magasra nőtt, amikor a szárazföldi növények egyike nem ért volna még derékig sem az embernek – már, ha lett volna akkor ember. Jóval a fa méretű növények megjelenése és elterjedése előtt, a szilur időszak végén és a korai devonban élt ez az élőlény, amely korának legnagyobb élő organizmusa volt, és igazi felhőkarcolóként tornyosult minden más korai szárazföldi növény fölé.
1843-ban a Kanadai Geológiai Intézet alapítója és első igazgatója, William Edmond Logan, akiről egyébként országának legmagasabb pontját, a Mount Logan-t is elnevezték, a québec-i Gaspé-öböl mentén végzett térképező és feltáró terepi munkát, hogy a Gaspé-félsziget szénben és egyéb ásványi anyagokban ígéretes képződményeit jobban megismerhesse a tudomány.
A munkálatok során a terület 420 millió éves, alsó-devon időszaki rétegeit is feldolgozták, és az ezekből a képződményekből származó növényi fosszíliák adták az egyik első, szisztematikusan tanulmányozható leletegyüttest a devon legkorábbi szakaszának flórájából. Az itt begyűjtött kövületek között volt egy érdekes, hatalmas fatörzsre emlékeztető képződmény, aminek átmérője akár egy méter is lehetett, egykori magassága pedig meghaladhatta a nyolc métert. A földtörténet későbbi korszakainak növényvilágában ez nem lenne nagy szenzáció, hiszen a fák többsége elérheti, vagy jócskán meg is haladhatja ezt a méretet. Azonban ne felejtsük el, hogy egy olyan korban járunk, amikor minden más növény ennél jóval kisebb méretű volt, az akkori flóra egyetlen eleme – a mohák, harasztok vagy korpafűfélék egyike sem haladta meg az egy méteres magasságot. Mi lehetett hát ez a saját korát jóval megelőző, torony magasságú kakukktojás a Gaspé-öbölből?
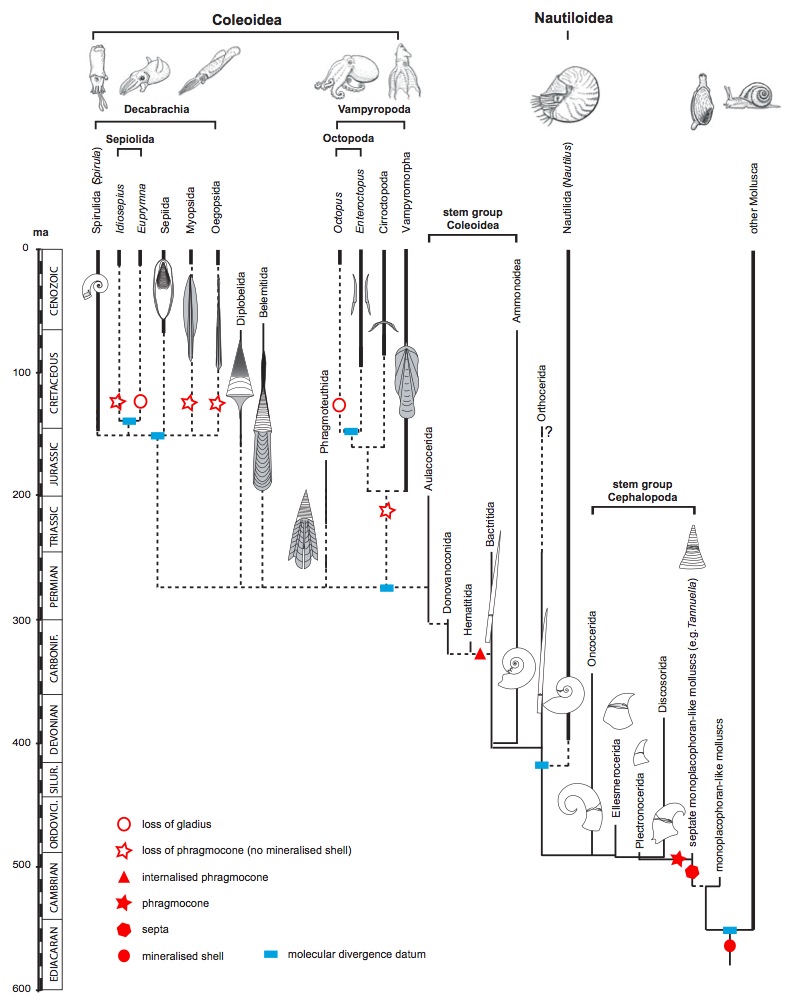
Érdekes módon a gyűjtemény ezek után hosszú évekig a polcokon porosodott, mígnem egy másik kanadai geológus, a későbbiekben a Darwin-féle evolúciós tanokat hevesen kritizáló műveiről is híres John William Dawson el nem végezte a különös kövület tudományos vizsgálatát. A már meglévő gyűjteményt saját maga által gyűjtött példányokkal is kiegészítette, majd végül 1859-ben keresztelte el a fosszilis fajt Prototaxites logani-nak. Védelmére szóljon, hogy a paleobotanika tudománya ebben az időben nem állt még a helyzet magaslatán, ráadásul Dawson a 7 napos teremtéselmélet elkötelezett híve is volt, így nem okozott neki problémát, hogy a hatalmas kövült törzseket egy egykori tobozos nyitvatermő fa maradványainak tekintse. A nemzetség általa adott neve, a Prototaxites is erre utal, ennek jelentése ugyanis ’első, vagy ősi tiszafa’, erre a mai növényre emlékeztették őt az ősmaradvány bélyegei. Ma már tudjuk, hogy a tobozos növények első képviselői valamikor a karbon időszak vége felé, 300 millió évvel ezelőtt alakultak ki, így a Prototaxites semmiképpen nem lehetett ennek a csoportnak a képviselője. Annyiban azonban már Dawson is közel állt az igazsághoz, miszerint a kövület bizonyos képletei alapján arra a következtetésre jutott, hogy annak bizonyos részét képezhették valamilyen gomba-szerű létformák maradványai is.
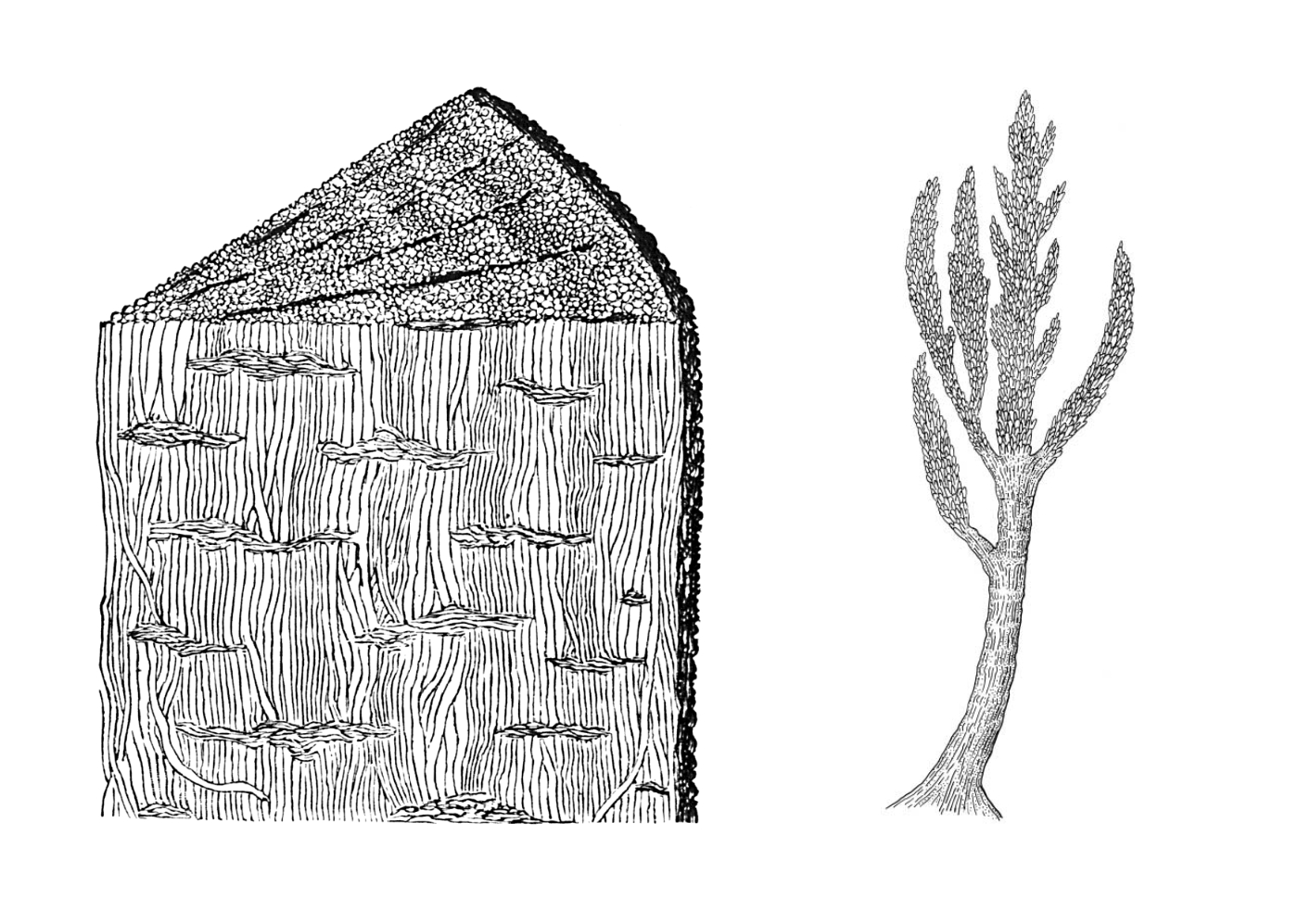 A Prototaxites metszeti rajza és egész alakos rekonstrukciója Dawson egy 1888-as tanulmányából.
A Prototaxites metszeti rajza és egész alakos rekonstrukciója Dawson egy 1888-as tanulmányából.
Ám nem sokáig volt egyeduralkodó a Prototaxites ezen interpretációja, 1872-ben ugyanis a Londoni Természettudományi Múzeum skót botanikusa, William Carruthers tanulmányában kifejtette, hogy Dawson ötlete a nyitvatermővel kapcsolatban egyenesen nevetséges, és – annak ellenére, hogy a kövület szárazföldi eredetére már ekkor is sok bizonyíték utalt – a titokzatos ősmaradványt a tengeri algák egy csoportjába sorolta. Lelkesedése odáig fajult, hogy Dawson eredeti elnevezését, a Prototaxites nemzetséget egyenesen felülírta saját ötletével, és a fosszíliának a Nematophycus, azaz a „szálas növény” nevet adta. Annak ellenére, hogy az ezt követő bő egy évszázad során pusztán egyetlen alkalommal próbálták cáfolni a rejtélyes, törzsszerű maradvány alga-mivoltát, a Nematophycus nevet végül hivatalosan nem fogadták el, és a tudományos nomenklatúrában a mai napig a szintén helytelen rendszertani besorolás nyomán megalkotott Prototaxites, ’ősi tiszafa’ név alatt találhatjuk meg ezt a fosszilis nemzetséget.
A fent említett egyetlen cáfolat még 1919-ben érkezett. Ekkor vetette fel egy szintén brit botanikus, Arthur Harry Church, hogy a Prototaxites felépítése és egykori élőhelyének ismert jellemzői akkor nyernek értelmet, ha azt feltételezzük, hogy valamiféle gomba volt. Sajnos az elgondolás – talán a háború utáni állapotok hatására – teljes apátiával szembesült a kor tudományos közegében, és a huszadik század hátralévő részében már nem is történt semmi érdemleges a Prototaxites kutatásának háza táján. Továbbra is kétséges maradt, hogy ez a hatalmas monstrum – amelynek példányai aztán a késő-szilurtól a késő-devonig terjedő időszak kőzeteiből több lelőhelyről is előkerültek – egy hínárszerű alga, egy méretes gomba vagy valamilyen zúzmóféle maradványa lehet-e.
A kutatás aztán az új évezred beköszöntével ismét fellendült.
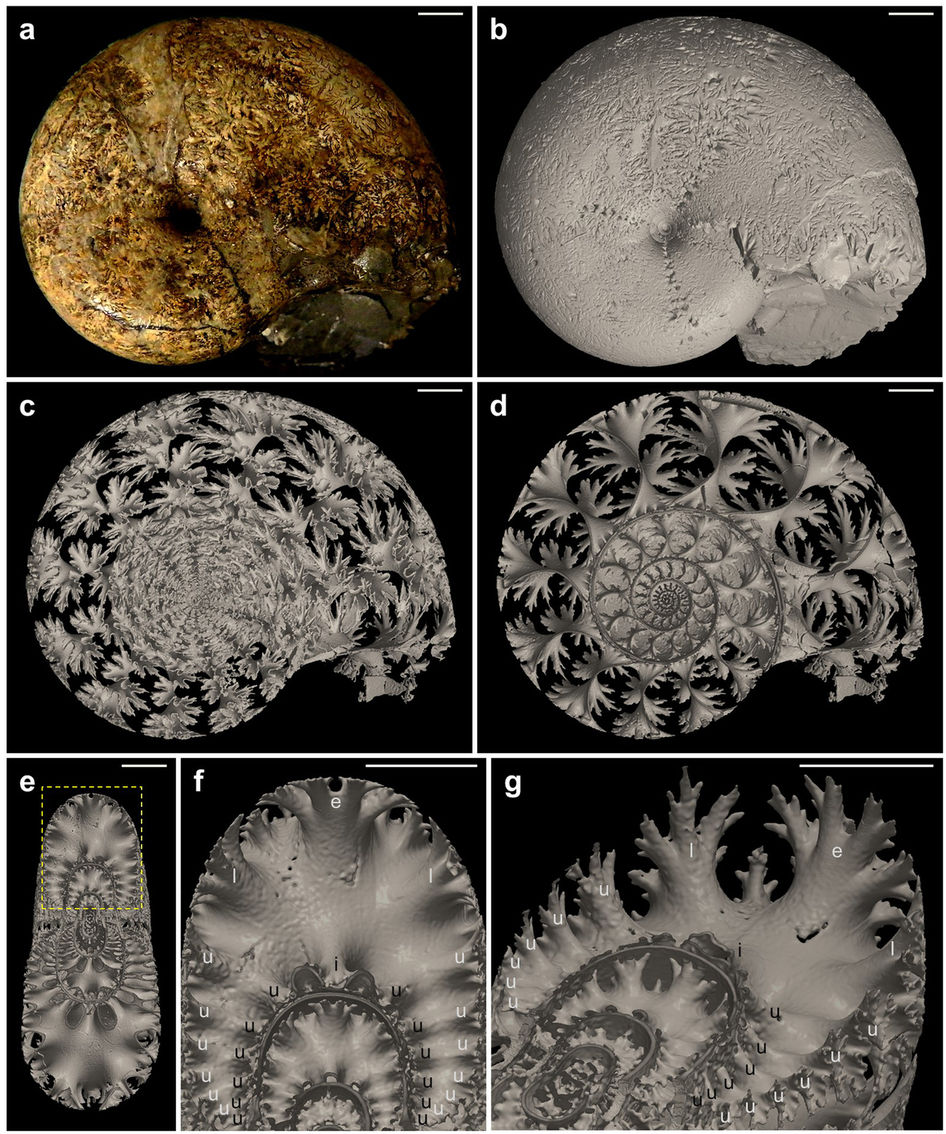
A washingtoni Smithsonian Intézet Természettudományi Múzeumának munkatársa, Francis Hueber – a fosszília keresztapjához, Dawsonhoz hasonlóan – saját gyűjtésű példányokat vizsgált, de ő nem csak Kanadából, hanem ausztrál és szaúd-arábiai lelőhelyekről is. Miután szorgalmasan többszáz vékony darabra vágta és csiszolta őket, Dawsonhoz hasonlóan ő is figyelmes lett a Prototaxites kövületeinek érdekes, nagyon apró, csőszerű képleteire, amelyek elektronmikroszkóp alatt úgy néztek ki, mint a mai gombák hifái, azaz gombafonalai. Ezek a gombafonalak micéliumokba rendeződve, az élőlény fő testéből kiindulva hatalmas hálózatot is létrehozhatnak, ami – a mai gombák egy részének esetében – más növényekkel való hasznos együttéléssel, szimbiózissal is járhat. Ennek egyik napjainkban sokat emlegetett változata a mikorrhiza.
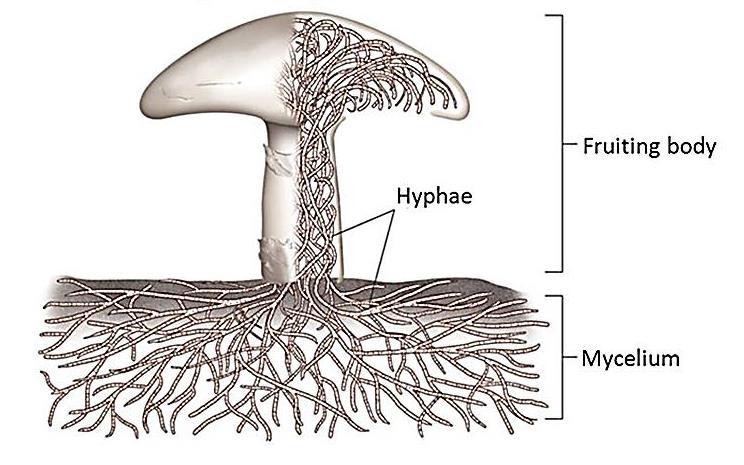 Egy mai gomba felépítése a gombafonalakkal (hyphae) és azok föld alatti hálózatával, a micéliummal (forrás: Katrin Tazelaar, Hogeschool INHolland)
Egy mai gomba felépítése a gombafonalakkal (hyphae) és azok föld alatti hálózatával, a micéliummal (forrás: Katrin Tazelaar, Hogeschool INHolland)
2001-ben megjelent tanulmányában az alaktani jegyek alapján tehát Hueber arra a következtetésre jutott, hogy a Prototaxites gomba volt. A későbbiekben ezt a tanulmányt újabb kutatások követték, és ezúttal a morfológia mellett a modern biogeokémiai ismereteket is segítségül hívták. Egy 2007-ben publikált vizsgálat során a maradványok szénizotópos összetételét vizsgálták, és jutottak a 2001-essel azonos eredményre. A kutatás alapját az képezte, hogy a növényekben – mivel minden esetben ugyanonnan, a légkör CO2 tartalmából nyerik a szenet – javarészt azonos tömegszámú izotópok találhatók, mint pl. 12C és 13C. Ha egy létforma nem fotoszintetizál, hanem heterotróf módon táplálkozik, tehát szerves anyagokat használ a túléléséhez szükséges kémiai energia biztosításához – például az állatok vagy a gombák –, a szenet az elfogyasztott szerves anyagból nyeri. Ebben az esetben a rendelkezésre álló táplálék függvényében változik a szénizotópok aránya is. Előszedték hát a lehető legtöbb, különféle lelőhelyekről és korokból származó Prototaxites fosszíliát, és részletes szénizotópos analízist végeztek rajtuk, figyelembe véve a későbbi, geológiai eredetű eltéréseket a görbéken. További kutatások aztán ezeket az adatokat – mintegy ellenőrzésképpen – összevetették olyan ma élő gombák eredményeivel is, amelyek a szilur végének kezdetleges, pionír társuláséihoz hasonló környezetben élnek. Az eredmény megfelelt a várakozásoknak: a szénizotópok jelentős diverzitást mutattak a különböző helyekről és időből származó példányok esetében, amely a változó környezeti tényezőkből adódó eltérő táplálékforrásra, szaprofita életmódra enged következtetni – és egyúttal arra, hogy a Prototaxites gomba volt!
 Egy hatalmas Prototaxites példány fosszíliája Szaúd-Arábiában (Forrás: Francis Hueber, 2001)
Egy hatalmas Prototaxites példány fosszíliája Szaúd-Arábiában (Forrás: Francis Hueber, 2001)
A vita azonban a mai napig nincs véglegesen lezárva. 2007 után több új munka született a Prototaxites rendszertani besorolását illetően. 2010-ben Linda E. Graham és társai egy komoly tanulmányban igyekeztek bizonyítani, hogy nem gombáról van szó, hanem a partmenti sekélytenger hullámtöréseitől, vagy más természeti erőktől szőnyeg módjára feltekeredett májmoha telepekről, olyasféle növényekről, mint amilyen a mai Marchantia. Ennek a feltételezésnek az alapját egyrészről a Prototaxites keresztmetszeteiben látható aszimmetrikus, évgyűrűhöz hasonló alakzatok, másrészről újabb szénizotópos vizsgálatok képezik. Ezt aztán még abban az évben hitelt érdemlően cáfolta egy másik kutatócsoport, érvelve a korábbi egyértelműen heterotróf életmódra utaló jelekkel, a szénizotópos vizsgálat módszertani hibáival és az új elképzelést teljesen felülíró, ám a „májmoha-elmélet” szerzői által elhallgatott anatómiai bélyegekkel. Egyúttal óvva intenek attól, hogy a Prototaxiteshez hasonló, kétséges rendszertani helyzetű és a távoli múlt ködébe vesző élőlényekkel kapcsolatban mindenáron jól beazonosítható mai analógiákat keressenek a kutatók.
 Kandis Elliot illusztrációja a Prototaxitesről mint különféle környezeti tényezők hatására szőnyegként feltekeredett májmoha telepekről - Linda Graham és társai elképzelése alapján.
Kandis Elliot illusztrációja a Prototaxitesről mint különféle környezeti tényezők hatására szőnyegként feltekeredett májmoha telepekről - Linda Graham és társai elképzelése alapján.
Legutóbb pedig 2014-ben jelentkezett egy kutatócsoport Retallack & Lanting vezetésével, akiknek az "óriásgomba-elmélettel" kapcsolatban több ellenvetésük is volt. Amellett, hogy a gombákhoz spórák is tartoznak, azt pedig soha nem találtak a Prototaxites közelében, nem látták reálisnak egy ekkora testű heterotróf, azaz elhullott élőlények maradványaival táplálkozó organizmus létjogosultságát sem egy olyan korai, fejletlen ökoszisztémában, mint ami a szilur végén rendelkezésre állt. Elvégre egy olyan környezetben – a miniatűr korpafüvek világában – ahol minden táplálékként felhasználható élő anyag mennyisége pusztán töredéke az enni akaró organizmusénak, valóban nehéz lett volna ilyen termetesre nőni. Ők úgy gondolják, hogy a megoldás az lehet, ha a Prototaxitest egyfajta korai zuzmóként értelmezzük. A zuzmók egysejtű fonalas kék- vagy zöldmoszatok és bizonyos gombafélék együttéléséből kialakult szervezetek, amelyek ily módon kétféleképpen is tudnak táplálkozni: a moszatok fotoszintetizálnak, míg a gombafonalak heterotróf anyagcserét végeznek.
Egy biztos: bármilyen élőlénycsoporthoz is tartozott, a Prototaxites korának legnagyobb élő szervezete volt, amely létezésének 70 millió éve alatt végigkísérte a földi növényvilág kibontakozását. Amikor 430 millió éve megjelent az élet színpadán, a növények még csak kezdetleges létformaként voltak jelen a szárazföldön, míg eltűnésükkor, 360 millió évvel ezelőtt már kifejlett erdőségek, és méretükben a Prototaxitesszel versengő fák uralták a bolygót.
Valószínű, hogy kihalásukért is az egyre fejlettebb szárazföldi állat- és növényvilág lehetett a felelős. A kezdeti időkben a Prototaxites tornyait semmi nem akadályozta abban, hogy szép lassan óriássá nőjenek, de ez szép lassan a nővények előretörésével megváltozott, és bár a rendelkezésükre álló tápanyag egyre több volt, az erdők szukcessziója végleg legyűrhette őket.
***
Ha tetszett a cikk, iratkozz fel hírlevelünkre, hogy ne maradj le az új tartalmakról!
Források:
- Hueber, F.M. (2001). "Rotted wood-alga-fungus: the history and life of Prototaxites Dawson 1859". Review of Palaeobotany and Palynology. 116 (1): 123–158. doi:10.1016/S0034-6667(01)00058-6
- Boyce, K.C.; Hotton, C.L.; Fogel, M.L.; Cody, G.D.; Hazen, R.M.; Knoll, A.H.; Hueber, F.M. (May 2007). "Devonian landscape heterogeneity recorded by a giant fungus" (PDF). Geology. 35 (5): 399–402. Bibcode:2007Geo....35..399B. doi:10.1130/G23384A.1.
- A Chicagoi Egyetem sajtóközleménye a 2007-es tanulmányról
- Graham, LE, Cook, ME, Hanson, DT, Pigg, KB and Graham, JM (2010). "Structural, physiological, and stable carbon isotopic evidence that the enigmatic Paleozoic fossil Prototaxites formed from rolled liverwort mats". American Journal of Botany. 97 (2): 268–275. doi:10.3732/ajb.0900322. PMID 21622387
- Taylor, T. N.; Taylor, E. L.; Decombeix, A. -L.; Schwendemann, A.; Serbet, R.; Escapa, I.; Krings, M. (2010). "The enigmatic Devonian fossil Prototaxites is not a rolled-up liverwort mat: Comment on the paper by Graham et al. (AJB 97: 268-275)". American Journal of Botany. 97 (7): 1074. doi:10.3732/ajb.1000047
- Retallack G.J. & Landing, E. (2014). "Affinities and architecture of Devonian trunks of Prototaxites loganii". Mycologia. 106: 1143–1156. doi:10.3852/13-390. PMID 24990121
- Paul Kenrick & Peter R. Crane (1997): The origin and early evolution of plants on land. NATURE | VOL 389 | 4 SEPTEMBER 1997. Page 33-39.
- A British Geological Survey honlapja: J D Hooker Slide Collection
- Falcon-Lang, H. (2012). Fossil “treasure trove” found in British Geological Survey vaults. Geology Today, 28(1), 26–30.